
2026/05/27
TOPICS
【プレスリリース】銅イオンの蓄積が細胞の低温死を引き起こすことを発見 -臓器の低温保存技術への発展に期待-

甲南大学大学院自然科学研究科の久原篤教授、山城芹奈 博士後期課程院生(日本学術振興会特別研究員)、太田茜 特任研究准教授(研究当時)(JSTさきがけ研究者、甲南大学研究員)らの研究チームは、「銅イオンによる細胞死『カプロトーシス』が動物の低温死に関与すること」を線虫の解析から明らかにしました。
近年発見された新しい細胞死 注1)である「カプロトーシス(銅誘導性細胞死)注2)」は、細胞内への銅イオン蓄積によって引き起こされます。しかし、カプロトーシスが動物個体内でどのような役割を持つのかは、これまで明らかになっていませんでした。本研究では、モデル生物である「線虫C.エレガンス 注3)」を用い、カプロトーシスが低温環境下での生存に深く関与していることを明らかにしました。研究チームは、リソソーム 注4)における銅輸送に関与するタンパク質SLCR-46.1に変異を持つ線虫に着目しました。この変異体 注5)(slcr-46.1)では、低温下で咽頭筋 注6)に銅イオンが過剰に蓄積し、低温死することが分かりました。
さらに、slcr-46.1変異体でカプロトーシス関連遺伝子の機能を抑制すると、低温下での生存率が回復しました。また、銅イオンをキレート 注7)することで変異体の低温死が抑制され、野生株 注8)においてもカプロトーシスを阻害すると低温死が抑制されました。さらに、哺乳類細胞を通常より低温条件で培養すると、カプロトーシス関連遺伝子の発現量が増加することも確認されました。
本研究は、生体内での役割が未解明であったカプロトーシスが、動物の低温死に関与することを示した初めての報告です。今後、ヒトを含む動物の低温適応機構の理解を深めるとともに、細胞や臓器の低温保存技術や細胞死制御などの新たな研究の発展につながることが期待されます。
本研究の成果は、2026年5月27日に、英国科学雑誌『Nature Communications』のオンライン速報版で公開されました。
<研究内容の模式図>

<研究の背景と経緯>
温度は、生き物の生死に直結する重要な環境要因です。日本では近年、「低温による死者数」が「熱中症による死者数」を上回る年が続いており、低温が健康や生命に与える影響への関心が高まっています。しかし、「なぜ低温が細胞や体を死に至らせるのか」という仕組みについては、まだ十分に分かっていません。
一方で近年、「細胞死」と呼ばれる現象の研究が進み、細胞がどのように死ぬのかについて新しい仕組みが次々と見つかっています。その一つが、銅という金属に依存して引き起こされる細胞死「カプロトーシス(cuproptosis)」です。この現象は比較的最近発見されたもので、主に細胞レベルでの研究が進められてきましたが、実際の生物個体においてどのような役割を持つのかはほとんど分かっていませんでした。
本研究では、モデル生物である線虫「C. エレガンス」を用いて低温に強い・弱い個体を調べる中で(図1)、細胞内での銅輸送に関与するタンパク質が低温での生死に関わることを発見しました。この結果から、「カプロトーシスが線虫の低温死に関わっているのではないか」という新たな仮説を立て、研究を進めました。
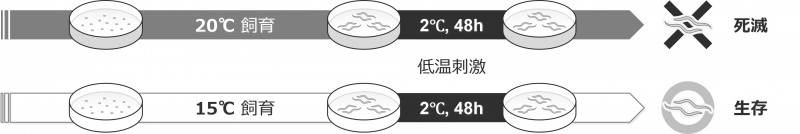
図1:線虫の低温耐性
通常、研究室でよく使われるイギリス・ブリストル由来の線虫「C. エレガンス」を20℃で育てた場合、2℃という低温に置かれると生きることができません。しかし、あらかじめ15℃で育てておくと、同じ2℃の環境でも生き残ることができます。これを「低温耐性 注9)」と呼びます(Ohnishi et al., Nat. Commun., 2024)。
<研究の内容>
低温耐性に関わる原因遺伝子の同定
低温耐性に異常を示す変異体を単離するため、線虫に化学変異原処理を行い、低温条件下で生存率が低下する個体をスクリーニング 注10)しました。その結果、低温耐性が著しく低下した変異体の単離に成功し、遺伝学的解析によって原因遺伝子を特定しました。
次世代DNAシーケンサー 注11)を用いた全ゲノム解析と染色体上での変異マッピングの結果、原因遺伝子は哺乳類SLC46A3の相同遺伝子 注12)であることが判明し、この遺伝子をslcr-46.1と命名しました。
slcr-46.1欠損変異体(以下、slcr-46.1変異体)では、低温下での生存率が有意に低下しました。一方、野生型のslcr-46.1遺伝子を導入すると、この表現型は回復しました。
リソソーム銅輸送体SLCR-46.1の機能解析
SLCR-46.1の発現細胞および細胞内局在を解析した結果、このタンパク質は咽頭筋や腸などで発現し、リソソーム膜に局在することが明らかとなりました。
さらに、slcr-46.1変異体の特定の組織で野生型のslcr-46.1遺伝子を発現させる解析から、slcr-46.1変異体における低温耐性の異常は、咽頭筋で野生型のslcr-46.1遺伝子を導入することで回復しました(図2)。これにより、咽頭筋におけるSLCR-46.1の発現が低温耐性に必須であることが示されました。

図2:slcr-46.1変異体における低温耐性テスト
野生株は通常、15℃で飼育した後に2℃に置かれても生存することができますが、slcr-46.1変異体は同じ条件下で死滅します。
この低温耐性の低下は、正常なslcr-46.1遺伝子全長を導入することで完全に回復しました。
slcr-46.1変異体の低温耐性の異常は、咽頭筋に正常なslcr-46.1遺伝子を導入することで回復しました。
哺乳類におけるslcr-46.1相同遺伝子の導入
slcr-46.1変異体で見られる低温耐性の異常は、咽頭筋に野生型のslcr-46.1遺伝子を導入することで回復しました。そこで、線虫slcr-46.1が哺乳類の相同遺伝子と共通した機能を持つのかを調べるため、マウスおよびヒトの相同遺伝子であるSLC46A3を線虫の咽頭筋で発現させました。その結果、マウスまたはヒトのSLC46A3をslcr-46.1変異体に導入すると、低温耐性の異常が回復することが分かりました(図3)。
これまでに、マウスSLC46A3は銅イオン輸送に関与する可能性が報告されており、この遺伝子を欠損させると細胞内の銅恒常性 注13)が乱れ、ミトコンドリア 注14)への異常な銅イオン蓄積が生じることが知られています。今回の結果は、線虫SLCR-46.1と哺乳類SLC46A3が共通した基質を輸送する可能性を示しています。
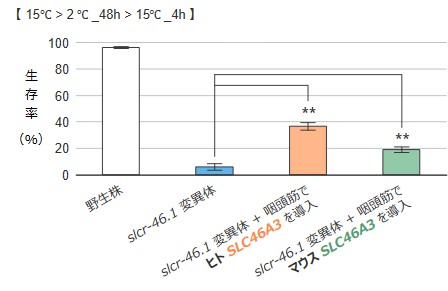
図3:slcr-46.1変異体に、哺乳類の相同遺伝子SLC46A3を導入した際の低温耐性テスト
線虫slcr-46.1変異体にヒトやマウスのホモログSLC46A3を導入すると、slcr-46.1変異体の低温耐性異常がいずれも部分的に回復しました。
低温刺激による銅イオン蓄積の可視化とその役割
銅イオン輸送への関与が示唆されているslcr-46.1のマウス相同遺伝子を導入すると、slcr-46.1変異体の低温耐性の異常が回復したことから、slcr-46.1変異体では野生株よりも銅イオンが異常に蓄積している可能性を検討しました。
そこで、銅イオンに応答して蛍光を発するプローブ 注15)(CF4)を用いて、線虫体内の銅イオンを観察しました。その結果、slcr-46.1変異体では、咽頭筋において野生株よりも強い銅イオン蓄積が確認されました(図4)。さらに、野生株においても低温刺激によって体内の銅イオン分布が変化することが分かりました。
特に、低温曝露後に死亡したslcr-46.1変異体では、咽頭筋の後端領域に顕著な銅イオン蓄積が見られました。
これらの結果から、咽頭筋における銅イオンの過剰蓄積がslcr-46.1変異体の低温死に関与することが示唆されました。
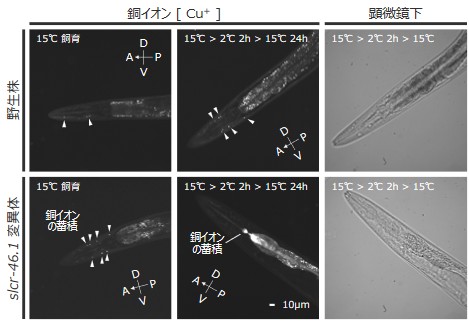
図4:低温刺激を与えられた線虫における銅イオンの蓄積
線虫を銅イオン蛍光プローブ(CF4)で染色し、体内の銅イオン分布を観察しました。
ショウジョウバエ細胞を用いたSLCR-46.1の機能解析
線虫slcr-46.1の細胞内での役割を詳しく調べるため、ショウジョウバエ由来の培養細胞 注16)(S2R+細胞)に線虫SLCR-46.1を導入し、その機能を解析しました。
SLCR-46.1に蛍光タンパク質(mCherry)を融合させてS2R+細胞に発現させたところ、SLCR-46.1はリソソーム膜に局在することが確認されました。
さらに、銅イオンに応答して蛍光を発するプローブ(CF4)を用いた解析により、SLCR-46.1を発現した細胞では、リソソーム内の銅イオン蓄積が増加していることが明らかになりました(図5)。
これらの結果から、SLCR-46.1はリソソーム内への銅イオン輸送に関与する可能性が示され、哺乳類ホモログであるSLC46A3と共通した機能を持つことが示唆されました。
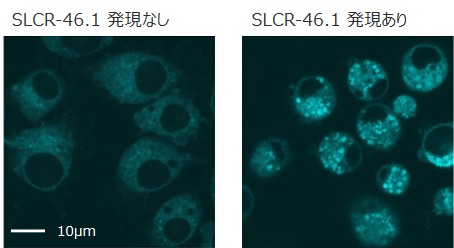
図5:S2R+培養細胞における銅イオンの可視化
ショウジョウバエ由来の培養細胞(S2R+細胞)を、銅イオン蛍光プローブ(CF4)で染色しました。線虫SLCR-46.1を発現した細胞では、銅イオン由来の蛍光シグナルが増加していることが確認できます。
カプロトーシス経路の関与
slcr-46.1変異体の咽頭筋において銅の過剰蓄積が見られたことから、銅誘導性細胞死「カプロトーシス」が低温死に関与するという仮説を立てました。この仮説を検証するため、カプロトーシス関連遺伝子であるfdx-2およびlias-1の機能をRNA干渉によって抑制しました。その結果、slcr-46.1変異体における低温耐性異常が回復しました(図6:左)。さらに、野生株においても、カプロトーシス関連遺伝子の機能を抑制すると低温耐性が向上しました(図6:中央、右)。
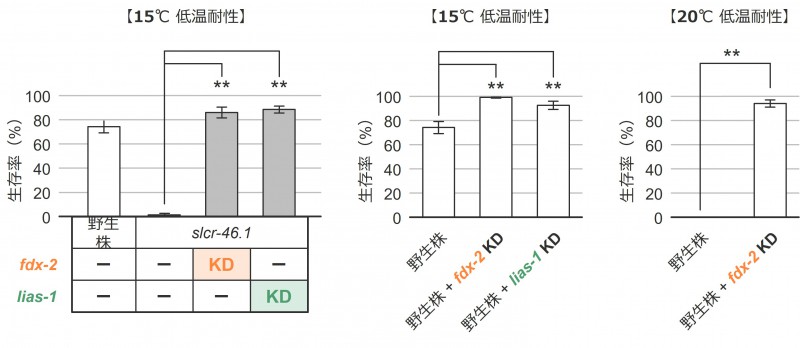
図6:カプロトーシス関連遺伝子の発現抑制(ノックダウン:KD)
カプロトーシスがslcr-46.1変異体の低温死に関与しているのかを調べるため、線虫におけるカプロトーシス関連の相同遺伝子(fdx-2、lias-1)の機能を、RNA干渉 注17)により抑制しました。
また、銅キレート剤「BCS」を用いてslcr-46.1変異体の細胞内銅濃度を低下させたところ、slcr-46.1変異体の低温耐性の異常が回復しました(図7)。これらの結果から、銅蓄積がslcr-46.1変異体の低温死の要因である可能性が示されました。
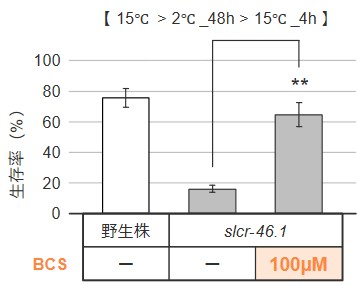
図7:銅キレート剤「BCS」による銅蓄積の阻害
線虫体内の銅蓄積がslcr-46.1変異体の低温死に関与しているかどうかを調べるため、銅キレート剤「BCS」を用いてslcr-46.1変異体の細胞内の銅濃度を低下させました。
ミトコンドリア機能への影響
また、低温刺激下のslcr-46.1変異体では、咽頭筋においてミトコンドリアの異常な集積が観察されました。このことから、低温死する線虫では、ミトコンドアの正常な代謝機能が損なわれている可能性が示されました(図8)。
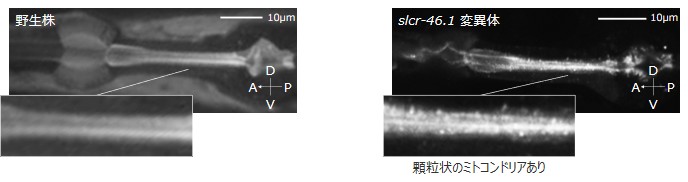
図8:低温刺激による、ミトコンドリアの異常集積
線虫をミトコンドリア蛍光プローブで染色し、体内のミトコンドリア分布を観察した。
細胞内銅代謝と低温ストレス応答の統合
以上の結果から、SLCR-46.1が関与するリソソームへの銅輸送が、低温時における細胞内の銅恒常性を制御し、その破綻によって咽頭筋における銅の異常蓄積とカプロトーシスが誘導されることが明らかとなりました。さらに、哺乳類細胞を通常より低温条件で培養すると、カプロトーシス関連遺伝子の発現量が増加することも確認されました(図9)。
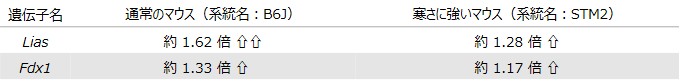
図9:低温下におけるカプロトーシス関連遺伝子の発現量の変化
(データソース: Suita et al., Cell Rep., 2023)
本研究により、細胞内金属代謝と細胞死経路が連動することで、個体レベルの低温ストレス応答が制御される新たな分子機構が示されました。
<研究の背景と経緯>
これまで、カプロトーシスが動物個体の低温死に関与することは知られていませんでした。そのため本研究は、「環境変化(寒さ)」と「銅による細胞死」を結びつける新たな視点を提示するものです。
今後、温度を感知する神経系と各組織における銅代謝や細胞死との関係を明らかにすることで、生物が個体レベルでどのように低温へ適応しているのかを統合的に理解できると期待されます。
また、哺乳類細胞においても低温環境下でカプロトーシス関連遺伝子の発現量が増加していたことから、本研究で見出した仕組みは種を超えて共通している可能性があります。
この発見は、ヒトの疾患との関連という観点からも注目されます。例えば、一部のがん細胞では銅が蓄積していることが知られており、カプロトーシスを人為的に誘導することで、新たながん治療法の開発につながる可能性があります。
さらに、低温下における細胞死制御の理解は、臓器移植や再生医療において重要となる細胞・組織の低温保存技術の向上にも貢献することが期待されます。
<研究支援>
本研究は中外製薬創薬科学財団、内藤記念科学振興財団、武田科学振興財団、日本医療研究開発機構(AMED)革新的先端研究開発支援事業(PRIME)「温度ストレスに対する微少金属イオンによる耐性制御」、G7財団、ホーユー科学奨励財団、旭硝子財団、JSPS科研費[JP22H04925、24H01255、26H01026、24K02070、25K22496、24K09538、24KJ2171]、JSPS(JP1270368)、JST PRESTO(25136813)、NIH(GM 79465)、東レ科学振興会の研究費によって行われました。
<用語解説>
注1)細胞死
細胞が自ら機能を停止し、死に至る現象のこと。細胞死には、不要になった細胞を除去する「アポトーシス」や、炎症を伴う細胞死など、さまざまな種類が存在する。近年では、特定の金属や代謝異常によって誘導される新しい細胞死機構も次々と発見されている。
注2)カプロトーシス(銅誘導性細胞死)
細胞内に銅イオンが過剰に蓄積することで誘導される新しいタイプの細胞死。2022年に報告された比較的新しい概念で、ミトコンドリア内の代謝異常を引き起こすことで細胞死に至ると考えられている。英語では「Cuproptosis」と呼ばれる。
注3)C.エレガンス
正式名称は「カエノラブディティス・エレガンス」で、土壌中に生息する非寄生性の線虫である。本種は分子遺伝学の分野で広く用いられており、1998年には多細胞生物として初めて全ゲノムDNAの解読が完了した。約2万個の遺伝子を持ち、その多くがヒトと共通していることから、ヒトの生命現象や疾患の理解に役立つモデル生物である。
注4)リソソーム・リソソーム膜
リソソームは、細胞内に存在する小器官の一つで、不要になったタンパク質や異物を分解・再利用する役割を持つ。細胞内の「リサイクルセンター」とも呼ばれ、近年では金属イオンの貯蔵や輸送にも関与することが分かっている。リソソーム膜は、リソソームを包み込む膜構造のこと。リソソーム内部の酸性環境や分解酵素を外部から隔離し、物質輸送を制御する役割を持つ。
注5)変異体
遺伝子に変化(変異)が導入された個体のこと。特定の遺伝子の機能を調べるために研究で広く用いられている。
注6)咽頭筋
線虫の口から腸へ食物を送り込むために働く筋肉組織。ポンプのように収縮することで細菌などを体内へ取り込む役割を持つ。本研究では、この組織で銅イオンが異常に蓄積していた。
注7)キレート(キレート剤)
金属イオンを強く結合して取り除く化学的作用、またはその働きを持つ物質のこと。銅キレート剤は細胞内の余分な銅イオンを捕捉し、銅による毒性を低下させるために用いられる。
注8)野生型・野生株
野生型とは、人為的な変異を導入していない、本来の正常な遺伝子や個体のこと。野生株とは、そのような遺伝的背景を持つ標準的な実験系統を指す。
注9)低温耐性
生物が低温環境下でも生存できる能力のこと。本研究では、線虫をあらかじめ低めの温度で飼育することで、通常は致死的な低温でも生存できるようになる現象を指す。
注10)スクリーニング
多数の個体や条件の中から、特定の性質(本研究では低温耐性の異常)を示すものを選び出す実験手法である。遺伝学では、目的の表現型を持つ変異体を効率的に見つけるために用いられる。
注11)次世代DNAシーケンサー
DNAの塩基配列を高速かつ網羅的に解析する装置である。従来法と比べて大量の配列情報を短時間で取得でき、全ゲノム解析や変異の同定に広く用いられている。
注12)相同遺伝子(ホモログ)
異なる生物種の間で共通の祖先に由来し、配列や機能が類似している遺伝子のことを指す。本研究では、線虫の遺伝子がヒトのSLC46A3と類似していることを示す。
注13)銅恒常性
細胞や生体内で銅イオン濃度を適切な範囲に維持する仕組みのこと。銅は生命活動に必須だが、過剰になると毒性を示すため、厳密な制御が必要である。
注14)ミトコンドリア
細胞内でエネルギーを生み出す働きを担う小器官。「細胞の発電所」とも呼ばれる。エネルギー代謝だけでなく、細胞死やストレス応答にも深く関与している。
注15)蛍光プローブ(CF4)
特定の分子やイオンに結合すると蛍光を発する試薬のこと。本研究で用いたCF4は銅イオンに反応して光るため、細胞内の銅分布を可視化することができる。
注16)培養細胞
生体から取り出し、人工的な環境下で増殖・維持されている細胞のこと。細胞レベルでの機能解析や薬剤評価などに広く用いられる。
注17)RNA干渉(RNAi)
特定の遺伝子の働きを一時的に抑える実験手法。目的遺伝子に対応したRNAを導入することで、その遺伝子から作られるタンパク質の量を減少させる。
<掲載論文名>
“Cuproptosis inducers mediate cold lethality via SLCR-46.1 in C. elegans”
(カプロトーシス誘導因子は、C. elegansにおいてSLCR-46.1を介して低温死を引き起こす)
英国科学雑誌「Nature Communications」
DOI:10.1038/s41467-026-73498-y
<発表者> *責任著者
山城芹奈(甲南大学大学院自然科学研究科 博士後期課程生、日本学術振興会特別研究員DC)
水野賢美(甲南大学大学院自然科学研究科 修士課程生(在籍時))
本村遥香(甲南大学大学院自然科学研究科 博士後期課程生、日本学術振興会特別研究員DC(在籍時))
Aidan T. Pezacki(米国プリンストン大学化学科 博士研究員)
森雪永(甲南大学大学院自然科学研究科 博士後期課程生、JST-Spring)
Christopher J. Chang(米国プリンストン大学化学科 教授)
*太田茜(甲南大学理工学部 研究員、JSTさきがけ研究者、特任研究准教授(研究当時))
*久原篤(甲南大学理工学部 大学院自然科学研究科 統合ニューロバイオロジー研究所 教授)